
 JA
JA
 EN
EN
 中文
中文




アレイ比較ゲノムハイブリダイゼーションを用いたねこなき症候群における遺伝子型‐表現型関係の高解像度マッピング論文の翻訳(ヒロクリニックNIPT仮訳)
1 カリフォルニア大学サンフランシスコ校総合がんセンター・実験室医学科、2 カリフォルニア大学サンフランシスコ校がん研究所、3 オーフス大学人間遺伝学研究所(デンマーク・オーフス)、4 コペンハーゲン大学医用生化学・遺伝学研究所(コペンハーゲン)、5 北京市中国科学院北京ゲノム研究所
はじめに
染色体5pの欠失はさまざまな発達上の欠損をもたらし、ほとんどの症例はねこなき症候群(MIM 123450)(Niebuhr 1978a)に分類される。 これらの欠失は末端または間質性であり、時に細胞遺伝学的に複雑な核型との関連で起こる(Sreekantaiah et al. 1999). ねこなき症候群には、その症候群の名前を与える特徴的な泣き声、顔面異形、音声遅延、精神遅滞(MR)など、いくつかの表現型の要素がある。
いくつかの以前の研究では、5p上の欠失セグメントの範囲と表現型との関連が示されている(Overhauser et al. 1994; Church et al. 1997; Marinescu et al. 1999a; Mainardi et al. 2001)。しかしながら、これらの研究は、削除された領域に対するMRの関係に関して、ある程度一貫性のない結果およびサブスタンス論争をもたらした。 異なる結果は、複数の観察者による表現型の一貫性のない評価、いくつかの表現型特性の顕著性の年齢依存性に対する考慮の欠如、および遺伝子型の評価に使用される分析技術の限界などの要因の組み合わせによる可能性が高い(Wilkinset al. 1983; Church et al. 1995; Marinescu et al. 1999a; Johnson et al. 2000)。本研究では、これらの潜在的問題の全てを取り上げた。
以前の遺伝子型解析では、従来の細胞遺伝学、FISH、多型マーカー、および最近では染色体ベースの比較ゲノムハイブリダイゼーション(CGH)が用いられてきた。(Marinescu et al. 1999b; Levy et al. 2002) 異常の性質を検出し、定義する。
従来の細胞遺伝学的解析は全ゲノムの概観を提供するが、解像度は限られている。 したがって、収差の境界は不正確に設定され、小さな欠失は見逃され、複雑な異常は不適切に同定される。 FISHおよび多型マーカーは、ゲノム配列に対する異常の正確な評価および異常のマッピングを可能にするが、プローブは個々にまたは少数のグループで評価されなければならないため、かなりの努力を必要とする。 したがって、多数の遺伝子座およびサンプルの研究は非常に労力を要する。 染色体CGHは、単一のハイブリダイゼーションでDNAコピー数変化の全ゲノムをスクリーニングすることができるが、その解像度は約5~10 Mbに制限され、結果はゲノム配列上に直接マッピングすることができない。
最近、著者らは、マイクロアレイCGHを実施する方法を開発した(Solinas-Toldo et al. 1997; Pinkel et al. 1998; Pollack et al. 1999; Snijders et al. 2001; Fiegler et al. 2003). 我々のアレイは、BACSやPACSなどの大型挿入ゲノムクローンから作製されたアレイエレメントを使用し、個々のクローンに影響する単一コピー異常の信頼性の高い検出を可能にするのに十分な測定精度を有する。 ゲノム配列上の各クローンの位置は、STSまたは末端配列などの配列タグを用いて決定される。 このようなアレイで得られる解像度は、クローン間のゲノム間隔およびクローン長によって決定される。 重複クローン(Albertson et al. 2000) を用いることにより、BACの長さの一部の解像度を得ることができる。
本研究では、アレイCGHを、通常の細胞遺伝学およびFISHを用いて5pに欠失を有すると決定された94人の患者のゲノムDNAの分析に適用した。 これらの標本は、我々の分析に先立って、規準に合致する詳細な表現型評価を受けた5p欠失症例の集積から選択した(Niebuhr 1978b;著者らの未発表結果)。 評価はほとんどの患者に対して数歳で実施し、表現型が最もよく評価される開発段階で確立できるようにした。 これらの症例のサブセットは、5p欠失の影響と遺伝子型-表現型相関の以前の研究に含まれている(Overhauser et al. 1990, 1994; Church et al. 1995, 1997). 著者らの結果は、これらの患者の評価のためのアレイCGHの価値を示し、ねこなき症候群におけるMR、音声遅延、顔面異形、および特徴的な泣き声の遺伝子型‐表現型相関を明らかにし、改良することを可能にした。
方法
ねこなき症候群の患者
被験者は、過去数十年間に我々の1人(E.N.)によって広範に研究された5p欠失患者150人以上のグループから選択された。 詳細な臨床データおよびゲノムDNAが入手できれば、被験者を組み入れた。 さらなる研究を容易にするために、著者らは、以前の研究が異常な染色体および異常な5番染色体との体細胞ハイブリッドの親起源に関する知識を提供し、家族性ゲノムDNAが利用可能であった被験者のみを分析した。 ほとんどの被験者について、従来の細胞遺伝学的分析の結果が利用可能であった。表1は、94人の適格な被験者をリストアップし、表現型評価、従来の細胞遺伝学的評価、およびアレイ結果を含む。患者番号1~69は、デンマークの連続した(診断時の)家族を指す。 100人以上の患者は、他国からの患者であり、ノルウェーからの患者100~117人、イギリスからの患者201~256人、チェコスロバキアからの患者300人、およびオーストラリアからの患者402人であり、より詳細な臨床的および細胞遺伝学的評価のために紹介された。 したがって、これらの症例は、5p欠失をもつ個体の準ランダムサンプリングである。 これらの検体のアレイCGH測定は、カリフォルニア大学サンフランシスコ校の治験審査委員会から適切な承認を得た後に実施した。
表現型の特徴を単一観察者(E.N.)により評価し、分類基準を一様に適用した。 表現型の側面は発達中に変化する可能性があるため、ほとんどの患者は5歳未満の場合と、5歳以上の場合に再度評価された。 表現型の特徴の有無は、表1において、それぞれ「Y」または「N」で標識される。 データの欠如は「ND」で示されており、表現型の特徴は年齢に依存している可能性があるため、いずれの年齢層の被験者にも「Y」が存在する場合は、その表現型の特徴が陽性であると分類される。 例えば、泣き声および顔面の形態異常は、患者が若いときに最も顕著であり、したがって、高齢群におけるこれらの表現型の喪失は有意であるとはみなされなかった。 いくつかの患者は、1つの年齢範囲で特定の表現型を示さず(「N」で示される)、他の患者ではデータが入手できず、その結果、表現型の状態に不確実性が生じた。 このような症例は、その特徴と遺伝子型との関係の決定から除外された。 したがって、患者18、45、56、112、および117は、クライ領域を定義するために使用されず、患者56および112は、顔面形態異常に関連するゲノム領域を決定するために使用されなかった。 最も重度の精神障害を有する患者は、他の理由で発話能力が障害されていた可能性があるため、発話遅延領域の確定には使用しなかった。 したがって、患者45は、表現型の音声成分の位置を特定するために使用されなかった。
MRの重症度を定義する精神運動発達評価は、個人的観察(E.N.)、文書情報、心理学的検査、学校成績情報などに基づいた。 MRは幼児では評価が困難であるため、欠失との相関には5歳以上のデータを有する患者のみを用いた。 したがって、22例、225例、231例は本解析から除外した。 MRの程度は、0(影響を受けていない)から7(重大な影響を受けている)の範囲の数値尺度によって示された。 各分類の一般的な判定基準を表2に示す。 これらの評価の不確実性は、尺度の下端で±0.5~上端で±1.0の範囲であると推定される。
アレイ-5pの高分解能分析のためのアレイには、遺伝的にマッピングされたSTSマーカーを用いて選択されたBAC、P1、およびPACクローンから作製されたエレメントが含まれていた(Hudson et al. 1995; Peterson et al. 1999)、および正常化およびその他の異常の検出に使用された他の染色体上に位置するクローンの可変数。 全てのクローンの位置は、ヒトゲノムの2003年7月のカリフォルニア大学サンタクルーズ(UCSC)凍結に基づいている。 5pの被覆密度は、ねこなき症候群で重要と以前に定義された領域で最も高かった。 これらの患者で認められた5p異常のすべてを包含する55クローンを表3に示す。ゲノム配列が完成すると、ほとんどのクローンを正確にマッピングすることができました。
しかし、いくつかの配列位置は遺伝地図情報に基づいて補間しなければならなかった。
全患者を、5p上に55クローン、および他の位置にさらに100~300クローンを含むアレイ上で分析した。 全ゲノムに分布する約1,750~2,000クローンを含む種々のバージョンのアレイについて、患者のうち37人を分析した(Snijders et al. 2001)、他の染色体が関与する異常の有無を調べる。 これらのより大きなアレイのいくつかは、5pクローンの完全なセットを含まなかった。
アレイエレメントのDNAをクローンから単離し、リンカー-アダプターPCRにより増幅した。 PCR産物を、約0.8μg/μlの濃度の水中20%ジメチルスルホキシドに懸濁した(Snijders et al. 2001年、キャピラリーチューブ印刷ピン(著者の未発表資料)を使用した特注プリンタを使用してクロム被覆スライド上にスポットした。 各クローンについて、隣接する3つのスポットを印刷した。 より小さいアレイについては、アレイ上の広く分離された位置で2セットの3連が印刷された。
アレイCGHハイブリダイゼーションおよび分析-患者および対照被験者の末梢血から、標準的な技術を用いて全ゲノムDNAを単離し、いずれかのニック翻訳(1μg)を用いてDNAを標識した(Pinkel et al. 1998)、またはランダムプライマー伸長(0.5μg)(Snijders et al. 2001). 標識にはdCTPに直接結合した蛍光色素を用いた。 いくつかの測定では、フルオレセインおよびテキサスレッド(またはAlexa 594)を使用したが、他の測定では、試験ゲノムおよび参照ゲノムをそれぞれ標識するためにCy3およびCy5を使用した。
反復配列のハイブリダイゼーションを抑制するために含まれる50μgのCot-1 DNAと共に標識試験および参照ゲノムDNAを、エタノール沈殿させ、ハイブリダイゼーションミックス中に再懸濁させて、50μlの全容積に対する50%ホルムアミド/10%硫酸デキストラン/2# SSC/1%-4%(v/v)SDSの最終組成にした。
ハイブリダイゼーション混合物を70℃に加熱してDNAを変性させ、温度を約1時間37℃に下げて、反復配列の再会合を可能にした。 ハイブリダイゼーションのために、アレイの周囲にゴムセメントから作られた低バリアを形成した。 次いで、ハイブリダイゼーション混合物をアレイに適用し、アレイを、流体の上面を自由にしたままのシールされたチャンバー内に置いた。 ハイブリダイゼーションは、37℃でゆっくりと揺動するテーブル上で16~48時間進行した(Pinkel et al. 1998).
ハイブリダイゼーション後、アレイを、ハイブリダイゼーション溶液の大部分を除去するために、PN緩衝液(0.1Mリン酸ナトリウム;0.1%ノニデットP40; pH 8)の流れで迅速に洗浄し、次いで、50%ホルムアミド/2# SSC中に45℃で15分間浸漬し、次いで、室温PN緩衝液中で15分間最終洗浄した。 アレイを洗浄した後、1μMのDNA染色DAPI(4′,6-ジアミジノ-2-フェニルインドール)を含有する90%グリセロールおよび10%リン酸緩衝液、pH 9を各アレイに適用し、カバースリップを加えた。 DAPIはアレイスポットを染色し、それらをハイブリダイゼーションシグナルとは無関係に可視化した。
アレイは、特注の電荷結合素子(CCD)イメージングシステム(Pinkel et al. 1998;著者の未発表データ)で画像化した。アレイ全体は1枚のCCD画像内に収まっていた。ほとんどの解析には、画像解析ソフトウェア UCSF SPOT (Jain et al. 2002)を使用した。このプログラムは、DAPI画像を使用してアレイのスポットの位置を決定し、局所的なバックグラウンドを差し引いた後、各スポット内の各ピクセルのテストおよびリファレンスの強度を計算した。各スポットの相対的なコピー数の指標として、(バックグラウンドを差し引いた)総テスト強度と(バックグラウンドを差し引いた)総参照強度の比を使用し、複製の比を平均化した。画像または比率データには、いかなる種類の計算上の調整(陰影補正、低さ正規化、空間依存的正規化、およびクローン特異的正規化)も行っていない。 問題信号を認識するために、スポット内の試験信号と基準信号の相関を含む品質基準を適用した。一般的に、相関 r < 0.8 のスポットは拒絶された。複製物についての SD >0.2 を有するクローンを分析から除去した。 ほとんどの場合、複製スポットのSDは<0.02であった。 また、品質検査に耐えた複製のうち1つのみを用いたクローンの測定も棄却した。 全体的な正規化係数を適用し、1セルあたり2コピーのアレイ要素のlog2比の中央値、すなわち中央値線形比を、それぞれ0または1に設定した。
図1

表1 ねこなき症候群患者94例の主要データの要約
| 患者 | 表現型 | レベル | 従来の細胞遺伝学a | 境界 (Mb/clone no.b) |
|||
|---|---|---|---|---|---|---|---|
| 特徴的な泣き声c | 顔面異形c | 音声遅延 | 近位d | 遠位 | |||
| 1 | Y/Y | Y/Y | Y | 5 | 46,XY,del(5)(p14.3)* | 20.837/39 | |
| 3 | Y/Y | Y/Y | Y | 6.5 | 46,XY,del(5)(p13.3)* | 30.726/51 | |
| 4 | Y/Y | Y/Y | Y | 6 | 46,XY,der(5)t(5;22)(p14.2;p13)pat | 30.022/44 | |
| 5 | Y/Y | Y/Y | Y | 5.5 | 46,XY,del(5)(p14.3)* | 17.500/37 | |
| 6 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p14.1)* | 23.276/41 | |
| 8 | Y/Y | Y/Y | Y | 6 | 46,XY,del(5)(p14.1)* | 25.841/42 | |
| 9 | Y/Y | Y/Y | Y | 7 | 45,XX,dic(5;13)(p13.3;p12)* | 31.942/45 | |
| 10 | Y/Y | Y/Y | Y | 5.5 | 46,XX,del(5)(p14.2)* | 21.797/40 | |
| 11 | ND/Y | Y/Y | Y | 5.5 | 46,XX,del(5)(p14.1)* | 30.022/44 | |
| 12 | ND/Y | Y/Y | Y | 5.5 | 46,XX,del(5)(p14.3)* | 18.336/38 | |
| 13 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p13.3)* | 33.428/49 | |
| 14 | Y/Y | Y/Y | Y | 6 | 46,XY,del(5)(p13.3)* | 30.022/44 | |
| 15 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.3)* | 20.837/39 | |
| 16 | Y/Y | Y/N | Y | 6.5 | 46,XX,der(5)t(5;8)(p14.1;p22)* | 31.751/46 | |
| 17 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p14.1)* | 25.841/42 | |
| 18 | ND/N | Y/N | N | 6 | 46,XY,del(5)(p14.1p15.31)*IN | 30.022/44 | 8.021/15 |
| 20 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p14.1)* | 31.751/46 | |
| 21 | Y/Y | Y/Y | Y | 5.5 | 46,XX,del(5)(p14.3)* | 18.336/38 | |
| 22 | Y/ND | Y/ND | Y | NAe | 46,XX,add(5)(p14.3)* | 21.797/40 | |
| 23 | Y/Y | Y/Y | Y | 6 | 46,XY,del(5)(p14.1)* | 30.022/44 | |
| 24 | Y/Y | Y/N | Y | 7 | 46,XX,add(5)(p14.1)mat | 21.797/40 | |
| 25 | Y/Y | Y/N | Y | 7 | 46,XX,add(5)(p14.2)* | 18.336/38 | |
| 26 | Y/Y | Y/Y | Y | 6 | 46,XY,del(5)(p14.1)* | 31.751/46 | |
| 27 | Y/Y | Y/Y | Y | 6 | 46,XY,del(5)(p13.3)* | 33.428/49 | |
| 28 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.3)* | 20.837/39 | |
| 30 | Y/Y | Y/Y | Y | 5.5 | 46,XX,del(5)(p14.3)* | 20.837/39 | |
| 31 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p13.3)* | 31.942/45 | |
| 35 | Y/ND | Y/ND | Y | 6.5 | 45,XY,der(5)t(5;14)(p14.2;q12),-14,mat | 23.276/41 | |
| 36 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p14.1)* | 30.022/44 | |
| 37 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p14.1)* | 30.022/44 | |
| 38 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p15.1)* | 18.336/38 | |
| 39 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p14.1)* | 30.022/44 | |
| 40 | Y/Y | Y/Y | Y | 5 | 46,XY,del(5)(p15.1)* | 16.684/33 | |
| 42 | Y/Y | Y/ND | Y | 3 | 46,XX,del(5)(p15.2)* | 11.035/28 | |
| 44 | Y/Y | Y/ND | Y | 3 | 46,XX,del(5)(p15.2)* | 11.035/28 | 1.207/1 |
| 45 | ND/N | N/N | Y | 7 | 46,XY,der(14)t(5;14)(q11;p13) del(5)(p14.3p15.31)* | 17.500/37 | 14.929/30 |
| 48 | Y/N | Y/N | Y | 6.5 | 46,XY,add(5)(p14.2)* | 16.684/33 | |
| 49 | Y/Y | Y/Y | N | 5 | 46,XX,der(5) ins(15;5)(q22;p14.3p15.31)mat | 26.794/43 | 5.945/12 |
| 50 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p14.1)* | 31.942/45 | |
| 51 | N/N | N/N | N | 0 | 46,XX,del(5)(p13.3p14.1)mat | 31.942/45 | 18.336/38 |
| 52 | Y/Y | Y/Y | Y | 6.5 | 46,XY,del(5)(p13.3)* | 31.751/46 | |
| 54 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p15.1)* | 16.684/33 | |
| 55 | Y/Y | Y/Y | Y | 5.5 | 46,XX,der(5)t(5;13)(p14.3;p12)mat | 20.837/39 | |
| 56 | ND/N | ND/N | ND | 6 | 46,XY,der(5)t(5;7)(p14.1p21)? del(5)(p14.1p15.1)?del(7)* | 26.794/43 | 8.021/15 |
| 58 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p14.1)* | 30.022/44 | |
| 59 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.3)* | 23.276/41 | |
| 60 | Y/Y | Y/Y | Y | 5 | 46,XY,del(5)(p14.2)* | 20.837/39 | |
| 63 | Y/Y | Y/Y | Y | 5 | 46,XX,rec(5)del(5)(p14.2) inv(5)(p14.1p15.1)mat | 21.797/40 | |
| 66 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p14.2)* | 23.276/41 | |
| 68 | Y/Y | Y/Y | Y | 6 | 46,XY,del(5)(p14.1)* | 25.841/42 | |
| 101 | Y/Y | Y/ND | Y | 2.5 | 46,XX,del(5)(p15.2)* | 11.035/28 | |
| 102 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.2p15.33)* | 23.276/41 | 3.174/4 |
| 104 | Y/Y | Y/Y | Y | 6.5 | 46,XY,del(5)(p13.3)* | 32.162/48 | |
| 105 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.3)* | 17.224/36 | |
| 107 | Y/Y | Y/Y | Y | 6 | 46,XY,del(5)(p13.3)* | 33.428/49 | |
| 109 | Y/Y | Y/Y | Y | 5.5 | mos 46,XX,del(5)(p14.1)/del(5)(p15.2)* | 25.841/42 | |
| 110 | Y/Y | Y/Y | Y | 6.5 | 46,XX,del(5)(p13.3)* | 30.726/51 | |
| 111 | Y/Y | Y/Y | Y | 6 | 46,XY,del(5)(p14.1)* | 25.841/42 | |
| 112 | ND/N | ND/N | N | 1 | 46,XX,del(5)(p14.2p15.2)* | 23.276/41 | 11.435/27 |
| 113 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.1)* | 23.276/41 | |
| 114 | N/N | N/N | N | 2 | 46,XY,del(5)(p14.2p15.2)* | 23.276/41 | 11.360/26 |
| 115 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p15.2)* | 11.035/28 | |
| 117 | ND/N | N/N | Y | 3.5 | 46,XY,del(5)(p15.31)* | 9.288/18 | |
| 201 | Y/Y | Y/N | Y | 7 | 46,XY,add(5)(p14.3)* | 17.224/36 | |
| 202 | Y/Y | Y/ND | Y | 2.5 | 46,XY,del(5)(p15.2)* | 11.435/27 | |
| 203 | Y/Y | Y/Y | Y | 6 | 46,XY,del(5)(p13.3)* | 30.022/44 | |
| 205 | Y/Y | Y/Y | Y | 5 | 46,XY,del(5)(p14.2)* | 21.797/40 | |
| 206 | Y/Y | Y/ND | Y | 7 | 46,XX,add(5)(p14.3)pat | 18.336/38 | |
| 209 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p13.3)* | 31.751/46 | |
| 210 | Y/Y | Y/Y | Y | 7 | 46,XX,del(5)(p14.1)pat | 25.841/42 | |
| 212 | Y/Y | Y/Y | Y | 6.5 | 46,XY,del(5)(p14.1)* | 30.022/44 | |
| 213 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.2)* | 20.837/39 | |
| 214 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.2)* | 17.224/36 | |
| 215 | Y/Y | Y/N | Y | 7 | 46,XX,der(5)t(4;5)(q32;p14.3)* | 14.929/30 | |
| 216 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.2)* | 20.837/39 | |
| 218 | Y/Y | Y/Y | Y | 6.5 | 46,XX,add(5)(p14.3)pat | 17.500/37f | |
| 219 | Y/Y | Y/Y | Y | 6.5 | 46,XY,del(5)(p14.1)* | 25.841/42 | 1.207/1 |
| 221 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.2)* | 18.336/38 | |
| 222 | Y/Y | Y/Y | Y | 6 | 46,XY,del(5)(p14.1)* | 26.794/43 | |
| 223 | Y/Y | Y/Y | Y | 6 | 46,XX,del(5)(p13.3)* | 33.428/49 | |
| 225 | Y/ND | Y/ND | Y | 4g | 46,XY,del(5)(p14.3p15.31)* | 21.797/40 3.174/4 | |
| 228 | Y/Y | Y/Y | Y | 6 | 46,XY,add(5)(p15.2)pat | 11.035/28 | |
| 229 | Y/Y | Y/Y | Y | 7 | 45,XX,dic(5;22)(p13.2 p12)* | 33.428/49 | |
| 231 | Y/ND | Y/ND | Y | 3.5g | 46,XX,del(5)(p15.1)* | 15.679/31 | |
| 232 | Y/Y | Y/Y | Y | 6.5 | 46,XX,del(5)(p13.3)* | 31.942/45 | |
| 250 | Y/Y | Y/Y | Y | 5 | 46,XX,del(5)(p14.1)* | 26.794/43 | |
| 251 | Y/Y | Y/Y | Y | 6.5 | 46,XX,del(5)(p13.3)* | 30.022/44 | |
| 252 | N/N | N/N | N | 5 | 46,XY,del(5)(p15.1p15.31)* | 15.872/32 | 6.365/13 |
| 253 | Y/Y | Y/Y | Y | 5 | 46,XY,r(5)(14.3q35.3)* | 20.837/39 | |
| 254 | Y/Y | Y/Y | Y | 5 | 46,XY,del(5)(p14.3)* | 20.837/39 | |
| 255 | Y/Y | Y/Y | Y | 6.5 | 46,XY,add(5)(p14.2)* | 20.837/39 | |
| 256 | Y/Y | Y/Y | Y | 5.5 | 46,XX,del(5)(p13.3)* | 30.022/44 | |
| 300 | Y/Y | Y/Y | Y | 5 | 46,XY,del(5)(p14.1)* | 25.841/42 | |
| 402 | Y/Y | Y/ND | Y | 3 | 46,XY,del(5)(p15.2)* | 11.035/28 | |
結果
アレイCGH測定
5番染色体および全ゲノムスキャンの代表的なコピー数プロファイルを図1に示す。 単一コピーの欠失および均一サンプルにおけるゲインは、理想的には、それぞれ-1.0および+0.58のlog2比を有するであろう。
実際には、欠失の比率は-1~-0.7の範囲であり、おそらく高度に反復された配列からのシグナルの不完全な抑制、交差ハイブリダイゼーション、バックグラウンドレベルの不完全な定義などにより、理想からの逸脱があった。 コピー数に対する比率とノイズレベル(図1に示されている)とのこのほぼ定量的な関係により、-0.4に等しいlog2比に設定された単純な閾値を使用することにより、非常に高い信頼性で欠失を検出することができた。 ほとんどの場合、2つのアレイクローン間のインターバルの一方または両方の欠失境界を確立するのに十分な精度が、単一のハイブリダイゼーションにおいて達成された。 最初の測定にノイズが多すぎて欠失境界に隣接するクローンを同定できなかった場合、測定を繰り返した。
色素リバーサルによる測定は、再ハイブリダイゼーション以上の情報を提供しなかったため、実施しなかった。 ときに、標本DNAの再精製がデータの質を改善し、いくつかの標本中の未知の汚染物質がゲノムDNAを一貫して標識またはハイブリダイズする能力に影響することを示すことが見出された。
表1に、患者94例の欠失データ、従来の細胞遺伝学的分析、および表現型特性を要約する。 患者は、以前の番号付け(Niebuhr 1978a; 著者の未発表資料)に対応して、番号順にリストアップされている。
82人の被験者における欠失は末端であったが、12人では欠失は間質性であり、5pで合計106個の欠失境界であった。 以下に詳しく述べるように、被験者のうち15例では5p欠失に加えてコピー数異常が認められた。 これらの異常のうち3つは欠失境界に隣接する5p配列の重複であったが、他の異常は表4に示すように5p以外のゲノム領域を含んでいた。
著者らの測定に基づいて、興味深いDNA配列特徴の2つの徴候が見出された。 最初に、有意なブレークポイントクラスターが検出された。 これらの患者で認められた106個の欠失境界のすべては染色体の遠位約33 Mb内に生じ、その結果、メガ塩基当たり約3個の境界の平均密度がもたらされた。
94例のうち、9例は5p15.2のクローン26、27、および28の約0.5 Mb領域に遠位または近位の境界を有し、メガ塩基当たり約18の境界を生じた。
さらに、この領域のクローンが2003年7月の配列凍結で示されているように順序付けられていると、欠失境界は複雑に見え、境界に隣接するクローンの比率に振動がある。 クローンを並べ替えると、すべての場合において欠失境界の見かけ上の構造が単純化される。 表3では、クローンを改訂した順序で列挙するが、ゲノム配列の集合に基づいて塩基配列の位置を示す。 これらの結果は、異常形成を促進し、ゲノム配列の適切な集合を複雑にするいくつかの配列モチーフがこの領域に存在する可能性を示唆する。 あるいは、シークエンスアセンブリが正しければ、この領域の欠失境界のクラスターは、局所的なシークエンス特徴を反映する反復複素数コピー構造を有する。 より高い密度のクローン被覆率は、このクラスター領域のサイズを潜在的に狭め、5pにおける他の切断点クラスターを潜在的に明らかにするであろう。
われわれが発見した第2の配列特徴は、正常ヒトゲノムにおける5p上の配列重複領域であった。 58例(例えば、図1Aおよび1B)のデータは、クローン39が、周囲のクローンが明確に欠失されたとき、正常領域で予想されるものと欠失の中間の、log2比~
-0.4を有することを示した。 しかしながら、欠失がクローン51および52の領域を含むように近位に延長された場合、クローン39上の比率は期待値まで低下した。 このことは、クローン39の配列のかなりの部分が、クローン51と52の間で5p上で複製されたことを示唆しており、クローン39のみを含む欠失は、4コピーから3コピー(log2[3/4] ~-0.4)への変化を表す。
このBACをプローブとして用いたFISH解析により、この仮定が確認され、5pに2つの近接した信号が発見された。 ヒトゲノム配列の一部の凍結(2002年6月、2002年11月、2003年4月)により、この重複の存在が示され、このBAC中のマーカーが20.791 Mbおよび34.070 Mb(2003年4月)の両方に位置することが示された。
2003年7月の凍結は、重複を示すものではありません。 このタイプの大きな重複はゲノム中で頻繁に起こる(Lupski et al. 1996; Eichler 2001)
DNAコピー数に多型は認められなかった(Albertson and Pinkel 2003; Iafrate et al. 2004; Sebat et al. 2004)クローン39を含む試料がこのセットで見つかった。 この領域における可能性のある配列複雑性のさらなる示唆は、表3に記載されているように、配列集合がクローン51近傍の我々の欠失境界の最も単純な解釈によって示されるクローンの順序と一致しないという事実である。
欠失に対するMRの依存性
図2Aは、5pの欠失と患者のMR状態との関係を示しており、患者は遅延のレベルを増加させることによって順序付けられている。 この関係は複雑で、いくぶん当惑しているように見える。 欠失の程度が増加するにつれて、遅延の重症度が明らかに増加するが、いくつかの重要な例外がある。 例えば、5pに間質性欠失を有する患者の中には、影響を受けていないか、またはごくわずかに遅延している患者もいるが、同じ領域に小さな欠失を有する患者では、著しく遅延している患者もいる。
従来の細胞遺伝学的およびFISH解析はこれらの被験者に対して以前に実施されており、一部の患者における染色体異常は複雑であり、5番染色体に加えて染色体の再配列が関与することが示されていた。 したがって、ゲノムワイドデータを提供するアレイを用いて、37例の患者を分析した。 これらの症例は、一般的な傾向と比較した場合、5p欠失が小さすぎて遅延レベルを説明できないと思われるすべての症例に加え、傾向に適合するほぼ同数を加えたものであった。 重度の遅延およびわずかな5p欠失が認められた被験者はいずれも、追加的な増減が認められた(例:患者45例[図1Cに示す])。
特異的な表現型への影響は、これらの追加的な異常のいずれにも特異的に起因するものではなかった。 異常が認められた被験者の詳細を表4に示す。 これらの測定には、進化するクローン組成を有するアレイを使用したため、追加の収差の境界が定義される精度は可変である。 しかしながら、これらの異常は、典型的には、少なくともいくつかの連続した配列要素を含み、非罹患者におけるコピー数多型よりもかなり大きくなっていることは明らかである(Albertson and Pinkel 2003; Iafrate et al. 2004; Sebatet al. 2004).
これらの複雑な例のいくつかは、欠失境界の近位にあるいくつかの重複した物質を含む、第5染色体に影響するさらなる異常を含む。 図2Bは、5p欠失を有し、他に異常が検出されなかった全ての患者について、欠失とMRレベルとの関係を示す。 MRの重症度と欠失の大きさと位置との関係は、今やはるかに一貫している。
泣き声、発声遅延、顔の形態異常に影響を及ぼす染色体領域
泣き声、発声、および顔の特徴に影響する染色体領域は、以前に5pの遠位部分にマッピングされている(Overhauser et al. 1994; Gersh et al. 1995, 1997).
図3は、これらの遺伝子型と表現型の関係に関する情報を提供する欠失境界をもつ被験者のデータを示している。 それぞれの場合について、境界にあるクローンを示す。 特徴的な泣き声に影響する領域は、患者49および252における差により最も狭く定義され、いずれも間質性欠失を有する。 cryを有する患者49については、欠失はクローン12の近位から始まり、cryを有さない患者252については、欠失はクローン14までの配列を保持し得る。 このように、この表現型に関与する染色体領域は、これらのクローンの間の1.5Mb領域に位置する。
特徴的な顔の特徴に影響する領域は、患者117、202、および114によって最も狭く定義される。 患者117は、クローン17と18の間に始まる末端欠失を有し、顔面表現型を有さない。 したがって、重要な領域はクローン17の近位に位置しなければならない。なぜなら、この位置まで材料を保持することができるからである。 患者202は、より大きな末端欠失を有し、クローン26と27との間の境界を有し、表現型を有する。 これは、クローン17と27の間の2.4-Mb領域を、顔の特徴を担う領域と定義する。 患者114は、この領域の近位限界を支持する。 患者114は間質性欠失を有し、クローン26と27との間の遠位境界を有し、表現型を有さず、この表現型の重要な位置をクローン27の遠位に置く。 本発明者らは、この提案された臨界領域を含む欠失を有する患者252は、患者が表現型を有さないため、不一致であることに留意する。
発語遅延に関与する染色体領域は、間質性欠失を有する患者102人、225人、および49人のデータによって最もよく定義される。 表現型を有する患者102および225では、欠失はクローン4の近位にある。 表現型を持たない患者49では、染色体の遠位部分はクローン13まで保持され得る。 したがって、音声遅延はクローン4と13の間の3.2-Mb領域内の欠失によるものである。
考察
発生症候群における患者の遺伝子型と表現型との関係の確立は、症状の原因となる遺伝的メカニズムの発見に向けた第一歩であり、患者における疾患の臨床的管理の基礎を提供する。 ねこなき症候群を引き起こす染色体異常に関する多くの以前の研究は、図4に要約されているように、この関係の概要を確立している。 古典的表現型の異なる成分に別々に影響する染色体5p上のいくつかの領域が定義されており、5p15.2の1つはMRに強い影響を及ぼし、「cri du chat critical region」(CdCCR)と呼ばれ、他は特徴的な泣き声、言語遅延、顔の特徴に関連している。 しかし、MRと欠失の関係についての議論を含め、種々の研究間に有意な相違があった。 アレイCGHを用いて、慎重に表現型を評価した患者群におけるDNAコピー数変化の効率的な高解像度分析を行った本研究は、表現型の特異的側面に関するより正確な情報を提供し、議論の余地のある問題のいくつかを明らかにした。
欠失サイズとMRとの関係を確認することは困難であり、いくつかの研究では、欠失サイズが増加するにつれて、影響の重症度が漸進的に増加することが明らかにされている(Mainardi et al. 2001年には、このような関係は見出されなかった(Marinescu et al. 1999a).
我々の研究では、我々の測定(Niebuhr 1978a, 1978b; Overhauser et al)の何年も前に、単一観察者(E.N.)によって一貫した方法でMR状態を数値スケールで評価された一連の患者を分析した。 1990年、Kjaer and Niebuhr 1999年。
遅延の評価は患者の年齢に依存する可能性があるため、我々は5歳以上の場合に評価を実施した患者のみを対象とした。 すべての分析は、エプスタイン-バーウイルス形質転換細胞株よりも末梢血から単離されたDNAを用いたため、形質転換誘発性の異常は我々の結果を混乱させなかった。
図2Aに5pの欠失とMRレベルの関係を示す。 MRレベルが約3までの患者はMRレベル5以上の患者よりも欠失が小さいことは明らかであるが、欠失サイズに対する遅延の依存性は弱いようである。 例えば、MRレベル5以上では、異なる遅延レベルの間で欠失範囲がほぼ完全に重複しているため、レベル-5のMRを有する多くの患者はレベル-7のMRを有する患者と同様の5p欠失を有する。 さらに、いくつかの顕著な不一致がある。 患者112および114は約10Mbの間質性欠失を有し、MRレベルは1~2であるが、顕著に罹患した患者45は同じ領域ではるかに小さい間質性欠失を有する。
細胞遺伝学的解析により、ねこなき症候群の一部の患者では、ゲノムの他の領域が関与するコピー数異常を伴う複雑な染色体再構成が認められることが以前に示されている。 従って、著者らは、5p欠失が小さすぎて遅延レベルを説明できないと思われる患者全員を含む37人の患者に対して全ゲノムアレイCGHを行った。 表4に示すように、これらの患者のうち15例では、5p欠失に加えて異常が認められた。 著者らの結果は、著しく遅延した対象の大部分(14/22)が5p欠失に加えて異常を有することを示した。 われわれが検出した唯一の異常が5pの欠失であった被験者に注意を向けさせると、欠失の大きさと位置に対する遅延の依存性がはるかに明らかになる。図2Bは、遅延レベル3.5以上について、欠失サイズが増加するにつれて遅延が単調に増加することを示している。 この一般的な行動からのわずかな逸脱は、患者の遺伝的因子の修飾、MRレベルの決定の不確実性、またはゲノムの他の部位で検出されなかった小さな異常によるものであろう。 図2Bに示すように、MRと欠失との関係が明確であることは、患者の表現型評価の正確さを裏付けるものであることに留意する。なぜなら、偶然にMRが起こった可能性は極めて低いからである。 欠失境界が5pterから33 Mbよりも近位にある患者がいないことは、このような欠失が発生中に致死的であることを示している可能性がある。
アレイCGHは、標準的な細胞遺伝学よりもゲノムのより完全な評価を提供する。 全体として、被験者の約16%に複雑な異常が認められ、これまでの試験(Niebuhr 1978a; Mainardi et al.)の12%~13%よりもやや大きかった。 2001).
アレイCGH(例、患者206)で新たに用量異常が認められた被験者の多く(全員ではない)では、5番染色体および他の染色体を含む細胞遺伝学的に検出された構造異常が認められた(表1)。
逆に、染色体間細胞遺伝学的異常を有する一部の被験者(例、患者4および49)では、染色体5p以外にコピー数の変化は認められなかった。
従って、アレイCGHのより高い分解能および効率は、ねこなき症候群における用量異常の評価に重要な利点を有し、特に重度の遅延患者に対して有意な診断的利点を有するべきであり、そのうちの約3分の2は中等度の5p欠失および追加の異常を有する。
図2Bに示すように、MRが5p欠失に依存していることから、この表現型に異なる影響を与える3つの染色体領域が存在することが示唆される。 最初のMRIは、遅延レベル2.5以上の全患者の欠失に含まれる。 患者117および56の所見は、MRIが5p15.31のクローン15と18の間の1.2- Mb領域に限局している可能性があることを示しており、これは、他の文献で定義されているように、5p15.2のCdCCRの末端にある(Overhauser et al. 1994; Mainardi et al. 2001).
どのくらいの重要な遺伝子がMRIに含まれているかは不明であるが、そのサイズが限られていることから、非常に少数である可能性が示唆される。
他の2つの領域、MRIIおよびMRIIIは、図2Bに示すように、MRIのすぐ近位に位置する。 MRIIに限定された欠失は軽度の影響を及ぼすが、MRIIIのみに影響を及ぼす欠失(例、患者51)は、識別可能な表現型をもたらさない。 しかしながら、MRIを含む欠失を有する患者は、欠失がMRIIおよびMRIIIに及ぶにつれて、次第に重度の遅延を来している。 このように、MRIIおよびMRIII内の一連の遺伝子が、おそらくMRIにおける遺伝子の影響の修飾を介して、表現型に寄与しているようである。
これらの境界の定義は、もちろん、いくらかの不確実性にさらされ、欠失がMRにどのように影響するかについて、根底にあるモデルを採用するかどうかに依存する。 われわれのモデルは基本的にMRIに支配的な役割を割り当てており、MRIIおよびMRIIIにおけるさらなる異常は、表現型の重症度を劇的に増大させる働きをする。 MRIの遠位境界は、クローン15より遠位の遺伝子の欠失または保持が我々の患者のMRレベルに有意な影響を及ぼさないという事実によって設定される。 しかし、MRIの遠位領域に限局した欠失を有する我々のセットの患者は存在せず、絶対的に確認することはできない。 欠失がないことは、MR表現型の欠如による確認の欠如を示すかもしれないが、そのような個体は、ねこなき症候群の他の特徴を有することを期待するであろう。
MRIの近位境界を設定することは、より問題が多い。
以上、我々は、患者117の境界を用いて、データと一致する最小可能領域の選択を示した。 しかし、それほど重症ではなく、より重症の遅延レベルを有する他の被験者では、わずかに大きな欠失が認められ、クローン26~28の近くの切断点クラスター領域の境界が認められる。 従って、患者117はいくぶん異常であり、そのため、MRIは、クローン28に対してさらに2 Mbを遠位側に延長することができる。 この領域をより明確に定義するためには、十分に特徴づけられた表現型を有する多数の追加症例の分析が必要であろう。
MRIIおよびMRIIIに限定された欠失は、軽度の罹患または表現型が正常な個体をもたらす。 しかし、比較的正常な表現型は「脆弱」であるように見える。なぜなら、他の異常が重度の表現型をもたらすからである。
これらの追加的な異常には、上述のようなMRIの欠失、および/または第5染色体(患者25、48、104、201、および251の場合)または他の染色体(表4)上の他の部位での増減が含まれる。
例えば、著しく遅延した患者45は、5pにMRII内にわずかな欠失しかなく、これは非常に軽度の遅延を生じると予想される。 このように、6qの追加欠失(図1C)は、おそらく非常に重度のMR表現型をもたらす。 われわれは、MRIIおよびMRIIIの増加が表現型に影響する可能性があることに留意する。 MRレベル6.5の患者48は、遅延レベル4~5と一致するMRIIおよびMRIIIの一部を欠失している。 このように、MRIIおよびMRIIIの一部の重複(欠失境界における)は強い効果を有する。
MRIIIの欠失を有する個人における正常な表現型の脆弱性も、家系データによって示される。 例えば、患者51における欠失は、母親および祖父母に存在し、3つの全ては影響を受けない。 しかしながら、MRIIIの欠失を有する罹患していない両親は、欠失を遺伝した罹患小児を有する可能性があるという報告がある(Hand et al. 2000、Johnson et al. 2000)。
この表現型の変化は、正常なゲノムをもつ人に有害な影響を及ぼさない環境的または遺伝的修飾因子が、MRIII欠失をもつ人に重大な結果をもたらすことを示している可能性がある。 あるいは、罹患した小児は、これらの研究で用いられた技術では検出されなかったさらなる異常を有することがある。 高解像度全ゲノムアレイCGHはこのような異常を検出し、このような小児の遺伝子型状態を明らかにする可能性がある。
著者らの測定はまた、特徴的な泣き、顔の特徴、および音声遅延に影響する染色体5p上の領域の改善された局在化を提供した。 cry領域では、1つのクローンのみの欠失によって表現型を有する患者と持たない患者が区別された。 これにより、2つのフランキングクローン(それぞれマーカーD5S2054およびD5S676を含む)の間に、可能性のある重要な遺伝子がアレイ上に配置される。 従って、これら2つのケースの高解像度分析は、臨界領域により厳しい限界値を配置することを可能にするであろう。 複数アレイクローンがそれらの中に含まれているので、現在の症例の高誤差解消分析は、音声遅延または顔領域を実質的に精緻化しないであろう。 遺伝子型-表現型関係に関する我々の結果を、これまでの公表文献と比較した結果を図4に示す。
要約すると、遺伝子型と表現型の関係の理解を改善するためにアレイCGHを用いて、5p欠失を有する94人の患者における異常をマッピングした。 著者らは、患者のMRレベルに異なる影響を及ぼす染色体の3つの領域があることを示した。 これら3つの領域の全部または一部が関与する欠失は、ゲノム中の他の異常と相まって、完全なMR表現型を生じるように相互作用する。 最後に、著者らの高解像度データは、ねこなき症候群における典型的な泣き、顔の特徴、および言語遅延に関与する遺伝子の位置の精緻化を可能にした。
表2
MRの分類
| MRレベル | 説明(IQ) | 表現型 | |
|---|---|---|---|
| 児童 | 幼児 | ||
| 0 | 正常 | MRはありません | MRはありません |
| 1 | 境目 (<70) |
標準的な学校に長年通い、マイナー/メジャーなサポートを必要とする | 正常な発達のマイルストーン、最初の学校の年に明らかな軽度の遅延 |
| 2 | 非常に穏やか (<65) |
標準的な学校に数年間通い、大きなサポートを必要とし、簡単な読み書きと数学の能力がある。 | 最初の数年間は正常な発達のマイルストーン、2-3歳から明らかな軽度の遅延 |
| 3 | 穏やか (<50) |
長文も含めて全てを理解し、非常に簡単な読み書きと算数の能力を持っています。 | 発達のマイルストーンは数か月間遅れています。 |
| 4 | 中程度 (<35) |
ほとんどのことを理解し、小さな文章とたくさんのサインを使う。 | 発達のマイルストーンが数か月遅れる。1歳から明らかな遅れ |
| 5 | 重度 (<20) |
簡単な日常文と単語を理解し、2~3語の文章と多くのサインを使用し、歩く。 | 発達のマイルストーンが数か月から1年遅れる。MRは1歳前に明らかになる |
| 6 | 非常に深刻 (<10) |
いくつかの言葉を理解できる。通常は、支えられていれば、不安定に歩いている。言葉がないか、いくつかの言葉しかない。 | 発達のマイルストーンは数年遅れている;MRは6か月前に明らかになる |
| 7 | 深遠 | 軽度または無反応を示し、座ったり立ったりすることができます。 | 5歳になっても独りで座ることができない。 |
表3
5p解析用アレイクローン
| クローン数 | マーカー | 位置 | クローン名称 | |||
|---|---|---|---|---|---|---|
| 記号 | 名称 | 細胞遺伝学a | 配列(Mb)a | 遺伝的(cM)b | ||
| 1 | D5S1981 | AFMa217zh1 | 5p15.33 | 1.207 | 1 | CTC-326E20 |
| 2 | D5S2005 | AFMB002xc1 | 5p15.33 | 1.395 | 0 | RP11-94J21 |
| 3 | D5S1970 | AFMa183wh5 | 5p15.33 | 2.497 | 5 | CTB-116F8 |
| 4 | D5S417 | AFM205wh8 | 5p15.33 | 3.174 | 6 | RP11-20B3 |
| 5 | D5S1980 | AFMA217yh1 | 5p15.33 | 3.449 | 7 | RP11-82M24 |
| 6 | D5S675 | AFM336tc1 | 5p15.33 | 3.998 | 9 | RP11-103L11 |
| 7 | D5S1906 | WI-2725 | 5p15.33 | 4.258 | CTB-27O23 | |
| 8 | D5S405 | AFM154xa3 | 5p15.33 | 3.995 | 9 | RP11-227M19 |
| 9 | D5S406 | AFM154xg3 | 5p15.32 | 5.047 | 12 | RP11-58A5 |
| 10 | D5S1921 | WI-2897 | 5p15.32 | 5.200 | CTC-263B18 | |
| 11 | D5S464 | AFM112xe3 | 5p15.32 | 5.890 | 15 | CTC-248O4 |
| 12 | D5S2054 | AFMB355wb1 | 5p15.32 | 5.945 | RP11-53K22 | |
| 13 | D5S635 | AFM276yb9 | 5p15.32/15.31 | 6.365 | 16 | RP11-36H5 |
| 14 | D5S676 | AFM347yg9 | 5p15.31 | 7.492 | 18 | RP11-46O23 |
| 15 | D5S18c | 5p15.31 | 8.021 | CTB-16G3 | ||
| 16 | D5S1957 | AFMa124wg5 | 5p15.31 | 8.550 | 20 | CTB-27I9 |
| 17 | D5S208 | 5p15.31 | 9.039 | 21 | DPA-896A3 | |
| 18 | D5S74d | 5p15.31 | 9.288 | DPA-671G7 | ||
| 19 | D5S630 | AFM268zd9 | 5p15.31 | 9.614 | 21 | DPA-255H3 |
| 20 | D5S759 | AFM204ze1 | 5p15.31/15.2 | 10.100 | DPA-1398C5 | |
| 21 | D5S1850 | WI6722 | 5p15.2 | 10.328 | DPA-1104F2 | |
| 22 | D5S23e | 5p15.2 | 10.364 | DPA-1349G2 | ||
| 23 | AFM042xa11 | 5p15.2 | 10.403 | 23 | RP11-145B1 | |
| 24 | D5S2480 | WI7320 | 5p15.2 | 10.732 | DPA-941C2 | |
| 25 | D5S432 | AFM255xb9 | 5p15.2 | 10.746 | RP11-72C10 | |
| 26 | D5S117 | 5p15.2 | 11.360 | DPB-70G7 | ||
| 27 | D5S2887 | AFMa240xf9 | 5p15.2 | 11.435 | 25 | RP11-29N3 |
| 28 | D5S478
f |
AFM179xd10 | 5p15.2 | 11.035 | 24 | CTC-305H11 |
| 29 | D5S2081 | AFM347ta5 | 5p15.2 | 13.530 | 26 | CTC-305H17 |
| 30 | D5S1991 | AFMa282wa5 | 5p15.2/15.1 | 14.929 | 29 | RP11-5N8 |
| 31 | D5S1989 | AFMa247wc1 | 5p15.1 | 15.679 | 30 | RP11-135M13 |
| 32 | D5S1954 | AFMA114xf9 | 5p15.1 | 15.872 | 30 | RP11-261B20 |
| 33 | D5S1963 | AFMA140vd1 | 5p15.1 | 16.684 | 31 | RP11-269O14 |
| 34 | D5S416 | AFM205wh10 | 5p15.1 | 16.723 | 31 | RP11-260E18 |
| 35 | D5S2114 | AFMa090yh5 | 5p15.1 | 16.949 | 32 | CTB-28D11 |
| 36 | D5S486 | AFM206zc1 | 5p15.1 | 17.224 | 34 | CTB-33B3 |
| 37 | D5S2096 | AFM105xg1 | 5p15.1 | 17.500 | 35 | RP11-88L18 |
| 38 | WI-4804 | 5p15.1/14.3 | 18.336 | CTB-34B4 | ||
| 39 | D5S2419g | WI-10830 | 5p14.3 | 20.837 | CTC-253L1 | |
| 40 | D5S411 | AFM193xe11 | 5p14.3 | 21.797 | 39 | CTB-55P22 |
| 41 | D5S1868 | WI-9400 | 5p14.3/14.2 | 23.276 | CTB-115E13 | |
| 42 | D5S648 | AFM292yg5 | 5p14.1 | 25.841 | 42 | CTB-100A5 |
| 43 | D5S627 | AFM217ye1 | 5p14.1 | 26.794 | 43 | CTC-296N5 |
| 44 | D5S661/D5s2061 | AFMC011xb1 | 5p14/13 | 30.022 | 44 | CTB-161H9 |
| 45 | D5S477f | AFM177xb4 | 5p13.3 | 31.942 | 49 | |
| 46 | D5S1993 | AFMa286ya9 | 5p13.3 | 31.751 | 48 | CTB-13O21 |
| 47 | D5S1986 | AFMA238za9 | 5p13.3 | 31.776 | 49 | RP11-5N11 |
| 48 | D5S1996 | AFM297wa5 | 5p13.3 | 32.162 | 51 | RP11-67P13 |
| 49 | D5S651 | AFM302wd5 | 5p13.3 | 33.428 | 51 | CTC-221E3 |
| 50 | D5S2062 | AFM277yh9 | 5p13.3 | 33.806 | 55 | RP11-94E6 |
| 51 | D5S395f | AFM284vc1 | 5p13.3 | 30.726 | CTB-62F24 | |
| 52 | D5S1506 | GATA63c02 | 5p13.3 | 33.873 | CTB-38G24 | |
| 53 | D5S395 | AFM063yb6 | 5p13.2 | 35.843 | 57 | RP11-85N3 |
| 54 | D5S2025 | AFMb297za5 | 5p13.2 | 36.018 | 57 | CTB-107L17 |
| 55 | D5S1994 | AFMa286ze9 | 5p13.2 | 36.465 | 56 | CTB-18O17 |
表4 追加の異常
| 患者 | MR レベル |
細胞遺伝学的位置 | コピー数 | 上限 | 下限 | 大きさ (Mb) |
改訂核型a | ||
|---|---|---|---|---|---|---|---|---|---|
| 保持クローンb | 位置 (Mb)c |
保持クローンb | 位置 (Mb)c |
||||||
| 16 | 6.5 | 8pter-21.3 | 3 | pter | 0 | RP11-89M8 | 22.745 | 22.745 | 46,XX,der(5)t(5;8)(p14.1;p21) |
| 24 | 7 | 13q32.2-ter | 3 | RP11-40H10 | 93.26 | qter | 113.042 | 19.782 | 46,XX,der(5)t(5;13)(p14.1;q32.2) |
| 25 | 7 | 5q35.3-ter | 3 | RP11-125L2 | 173.496 | qter | 181.034 | 7.538 | 46,XX,der(5)t(5;5)(p14.2;q35.3)mat |
| 35 | 6.5 | 14q11.2 | 1 | RP11-152G22 | 19.227 | RP11-68M15 | 21.467 | 2.24 | 45,XY,der(5)t(5;14)(p14.2;q11.2), -14,mat |
| 45 | 7 | 6q25.2-6qter | 1 | GS-59B4 | 152.455 | RP1-57H24 | 170.695 | 18.24 | 46,XY,der(14)t(5;14)(q11;p13) del(5)(p15.1p15.2)del(6)(q25.2) |
| 48 | 6.5 | 5p15.11-5p14 | 3 | RP11-261B20/32d | 15.872 | CTC-296N5/43 | 26.794 | 10.922 | 46,XY,del(5)(p14.3)dup(5)(p14.2p15.1) |
| 104 | 6.5 | 5p13.3 | 4 | CTB-13O21/46 | 31.715 | RP11-67P13/48 | 32.162 | .447 | 46,XY,del(5)(p13.3)dup(5)(~p13.3)e×2 |
| 201 | 7 | 5q35.1-terf | 3 | RP11-15F10 | 169.122 | qter | 181.034 | 11.912 | 46,XY,der(5)t(5;5)(p14.3q35.1) |
| 206 | 7 | 20pter-12.2 | 3 | pter | 0 | DuPontA-967A8 | 10.282 | 10.282 | 46,XX,der(5)t(5;20)(p14.3;p12.2)pat |
| 210 | 7 | 1qter | 3 | RP11-194F13 | 238.741 | qter | 246.127 | 7.386 | 46,XX,der(5)t(1;5)(q43;p14.1)pat |
| 215 | 7 | 4q31.1-ter | 3 | RP11-40D5 | 135.249 | qter | 191.731 | 56.482 | 46,XX,der(5)t(4;5)( q31.1;p14.3) |
| 218 | 6.5 | 8pter-8p23.2 | 3 | pter | 0 | RP11-112O08 | 11 | 11 | 46,XX,der(5)t(5;8)(p14.3;p23.2)pat |
| 228 | 6 | 8q23.1-ter | 3 | CTD-2013D21 | 110.377 | qter | 146.308 | 35.931 | 46,XY,der(5)t(5;8)(p15.2;q23.1)pat |
| 251 | 6.5 | 5p13.3 | 3 | CTB-115E13/41 | 23.276 | RP11-94E6/50 | 33.806 | 10.53 | 46,XY,del(5)(p13.3)dup(5)(p13.3~14.1)e |
| 255 | 6.5 | 1qter | 3 | RP11-210E16g | 229.578 | 46,XY,der(5)t(1;5)(q42~43;15.2) | |||
図2
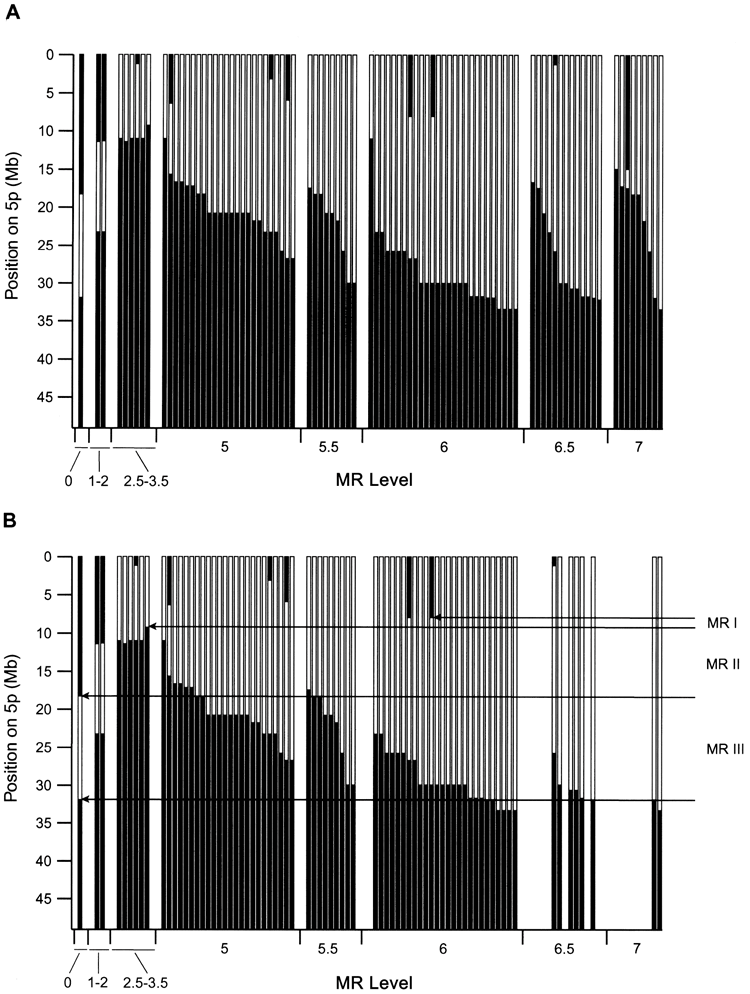
5p欠失に対するMRレベルの依存性。 A、遅延評価が入手できた全91例のデータ。 黒い部分は、それぞれの場合に保持されている染色体領域を示し、5pterからの物理的位置に対してプロットした。 セントロメアはグラフの下端のすぐ下にある。 欠失に対する遅延レベルの依存性は明白であるが、一般的な傾向には小さすぎるように見える欠失を伴う場合が多い。 MRレベル5については、各レベル内の近位欠失境界の順にデータがプロットされる。
これらの重度かつ重度の罹患患者からの遅延表現型のMRレベルの割り付けには、0.5~1の不確実性が推定されている。 5pの欠失のみを検出した患者についても、B、同じプロットを用いた。 重度の障害をもつ患者の大半は、さらに異常があるため、もはや存在しないことに注意。 5p欠失に対する遅延の依存性ははるかに一貫している。 5p‐MRI、MRII、およびMRIIIの3つの領域が、遅延に対する差次的効果を有することが示唆される。 これらの領域については本文で説明する。
図3 Am. J. Hum. Genet. 76:312–326, 2005

黒い四角は、その患者に関連する情報が入手できなかったことを示している。 丸印は、その特徴の原因となる染色体部分の位置に関して最も有益な情報を提供する患者を示す。 患者252の網掛けされた「N」は、この患者の欠失が、そうでないことによって表現型を生じさせたはずであることを示す。
図4 Zhang et al.: Cri du Chat Genotype-Phenotype

謝辞
本研究は、米国国立小児保健・人間開発研究所の助成金HD 17665、株式会社ヴァイシス、デンマークSTF比較ゲノムプログラム、デンマーク基礎研究基金の統合的生物学プラットフォーム、中国国立自然科学財団の支援を受けた。
電子データベース情報
- ここに掲載されているデータのURLは以下の通り。オンラインメンデル遺伝(OMIM)、http://www.ncbi.nlm.nih.gov/Omim/ (ねこなき症候群用)、UCSC ゲノムバイオインフォマティクス、http://genome.cse.ucsc.edu/ 参考文献Albertson DG、Pinkel D(2003)ヒト遺伝病およびがんにおけるゲノムマイクロアレイ。
参考文献
- Hum Mol Genet 12(Spec 2):R145-152 Albertson DG、Ylstra B、Segraves R、Collins C、Darkee SH、Kowbel D、Kuo WL、Gray JW、Pinkel D(2000)アレイCGHによるアンプリコン構造の定量的マッピングにより、CYP24が候補がん遺伝子であることが確認された。
- Nat Genet 25:144-146 Church DM, Bengtsson U, Nielsen KV, Wasmuth JJ, Niebuhr E (1995) 異なる表現型の特徴をもたらす遠位5pの異なるセグメントの欠失の分子的定義。
- Am J Hum Genet 56:1162-1172 Church DM, Yang J, Bocian M, Shiang R, Wasmuth JJ (1997) ヒト染色体5pのねこなき領域の高解像度物理および転写マップ。
- Genome Res 7:787-801 Eichler EE (2001) 最近の重複、ドメインの付加、およびヒトゲノムの動的突然変異。
- Trends Genet 17: 661-669 Fiegler H, Carr P, Douglas EJ, Burford DC, Hunt S, Scott CE, Smith J, Vetrie D, Gorman P, Tomlinson IP, Carter NP(2003) DNAマイクロアレイを用いて、BACおよびPACクローンのDOP-PCR増幅に基づく比較ゲノムハイブリダイゼーションを行った。
- Genes Chromosomes Cancer 36:361-374 Gersh M、Goodart SA、Pasztor LM、Harris DJ、Weiss L、Overhauser J (1995) 5p欠失を有する患者において猫のような鳴き声を引き起こす異なる領域に関する証拠。
- Am J Hum Genet 56: 1404-1410 Gersh M, Grady D, Rojas K, Lovett M, Moyzis R, Overhauser J (1997) 間期FISHを用いた5p欠失の分析のための診断ツールの開発
- Cytogenet Cell Genet 77: 246-251 Hand JL, Michels VV, Marinello MJ, Ketterling RP, Jalal SM (2000) 染色体5pおよび16qの間質性欠失が明らかな表現型効果なしに遺伝した: furth 確認。
- Prenat Diagn 20:144-148 Hudson TJ、Stein LD、Gerety SS、Ma J、Castle AB、 Silva J、 Silva J、 Slonim DK、Baptista R、Kruglyak L、Xu S-H、Hu X、Colbert AME、Rosenberg C、Reeve-Daly MP、Rozen S、Hui L、Wu X、Vestergaard C、Wilson KM、Bae JS、Maitra S、Ganiatsas S、Evans CA、DeAngelis MM、Ingals KA(1995) ヒトゲノムのSTSベース地図。
- Science 270:1945-1954 Iafrate AJ, Feuk L, Rivera MN, Listewnik ML, Donahoe PK, Qi Y, Scherer SW, Lee C (2004) ヒトゲノムにおける大規模変異の検出
- Nat Genet 36:949-951 Jain AN, Tokuyasu TA, Snijders AM, Segraves R, Albertson DG, Pinkel D (2002) マイクロアレイ画像データの完全自動定量化
- Genome Res 12:325-332 Johnson EI, Marinescu RC, Punnett HH, Tenenholz B, Overhauser J (2000) 5p14欠失は小頭症および発作に関連していた。
- J Med Genet 37:125-127 Kjaer I, Niebuhr E (1999) ねこなき症候群患者23例における頭蓋底の研究は、この状態に関与する頭蓋発達場を示唆している。
- Am J Med Genet 82:6-14 Levy B, Dunn TM, Kern JH, Hirschhorn K, Kardon NB (2002) dup5q/del 5p(cri du chat)患者における分子細胞遺伝学的解析によるdup5q表現型のDelineation
- Am J Med Genet 108:192-197 Lupski JR, Roth JR, Weinstock GM (1996) 細菌、ショウジョウバエ、ヒトにおける染色体重複。
- Am J Hum Genet 58:21-27 Mainardi PC, Perfumo C, Cali A, Coucourde G, Pastore G, Cavani S, Zara F, Overhauser J, Pierluigi M, Bricarelli FD (2001) 5p欠失患者80例の臨床的および分子的特徴: 遺伝子型-表現型相関
- J Med Genet 38:151-158 Marinescu RC, Johnson EI, Dykens EM, Hodapp RM, Overhauser J (1999a) 欠失の大きさとねこなき症候群における発達遅延のレベルとの間に関係はない。
- Am J Med Genet 86:66-70 Marinescu RC, Johnson EI, Grady D, Chen XN, Overhauser J (1999b) ねこなき症候群と診断された患者における末端欠失のFISH解析
- Clin Genet 56:282-288 Niebuhr E (1978a) 5p-核型を有する35例の細胞学的観察。
- Hum Genet 42:143-156 — (1978b) ねこなき症候群: 疫学、細胞遺伝学、および臨床的特徴。
- Hum Genet 44:227-275 Overhauser J, Huang X, Gersh M, Wilson W, McMahon J, Bengtsson U, Rojas K, Meyer M, Wasmuth JJ (1994) 第5染色体短腕の分子および表現型マッピング: ねこなき症候群の臨界領域の亜局在化。
- Hum Mol Genet 3:247-252 Overhauser J, McMahon J, Oberlender S, Carlin ME, Niebuhr E, Wasmuth JJ, Lee-Chen J (1990) ねこなき症候群における染色体5欠失の親起源。
- Am J Med Genet 37:83-86 Peterson ET, Sutherland R, Robinson DL, Chasteen L, Gersh M, Overhauser J, Deaven LL, Moyzis RK, Grady DL (1999) ヒト5番染色体短腕の統合物理地図。
- Genome Res 9:1250-1267 Pinkel D、Segraves R、Sudar D、Clark S、Poole I、Kowbel D、Collins C、Kuo WL、Chen C、Zhai Y、Dirkee SH、Ljung BM、Gray JW、Albertson DG(1998)マイクロアレイに対する比較ゲノムハイブリダイゼーションを用いたDNAコピー数変異の高分解能分析
- Nat Genet 20:207-211 Pollack JR, Perou CM, Alizadeh AA, Eisen MB, Pergamenschikov A, Williams CF, Jeffrey SS, Bo stein D, Brown PO (1999) cDNAマイクロアレイを用いたDNAコピー数変化のゲノムワイド分析
- Nat Genet 23:41-46 Sebat J、Lakshmi B、Troge J、Alexander J、Young J、Lundin P、Maner S、Massa H、Walker M、Chi M、Navin N、Lucito R、Healy J、Hicks J、Ye K、Reiner A、Gilliam TC、Trask B、Patterson N、Zetterberg A、Wigler M(2004)ヒトゲノムにおける大規模コピー数多型。
- Science 305:525-528 Snijders AM、Nowak N、Segraves R、Blackwood S、Brown N、Conroy J、Hamilton G、Hindle AK、Huey B、Kimura K、Law S、Myambo K、Palmer J、Ylstra B、Yue JP、Gray JW、Jain AN、Pinkel D、Albertson DG(2001)ゲノムワイドDNAコピー数測定用マイクロアレイのアセンブリ
- Nat Genet 29:263-264 Solinas-Toldo S, Lampel S, Stilgenbauer S, Nickolenko J, Benner A, Dohner H, Cremer T, Lichter P (1997) マトリックスベースの比較ゲノムハイブリダイゼーション: ゲノム不均衡をスクリーニングするためのバイオチップ。
- Genes Chromosomes Cancer 20:399-407 Sreekantaiah C, Kronn D, Marinescu RC, Goldin B, Overhauser J (1999) 典型的な猫のような泣き声があり、ねこなき症候群の他の臨床所見がない患者における複雑な染色体再構成の特徴づけ。
- Am J Med Genet 86:264-268 Wilkins LE, Brown JA, Nance WE, Wolf B (1983) ねこなき症候群の自宅で育てた小児80例における臨床的不均一性
- J Pediatr 102:528-533
出版社の注記
スプリンガー・ネイチャーは、公開されている地図や施設内の提携における管轄権の主張に関しては中立的である。








